- Record: found
- Abstract: found
- Article: found
Rice actin binding protein RMD controls crown root angle in response to external phosphate
Read this article at
Abstract
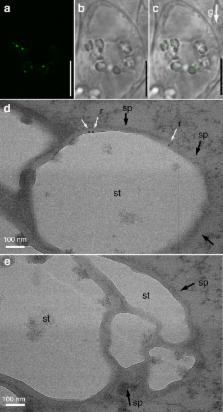
Root angle has a major impact on acquisition of nutrients like phosphate that accumulate in topsoil and in many species; low phosphate induces shallower root growth as an adaptive response. Identifying genes and mechanisms controlling root angle is therefore of paramount importance to plant breeding. Here we show that the actin-binding protein Rice Morphology Determinant (RMD) controls root growth angle by linking actin filaments and gravity-sensing organelles termed statoliths. RMD is upregulated in response to low external phosphate and mutants lacking of RMD have steeper crown root growth angles that are unresponsive to phosphate levels. RMD protein localizes to the surface of statoliths, and rmd mutants exhibit faster gravitropic response owing to more rapid statoliths movement. We conclude that adaptive changes to root angle in response to external phosphate availability are RMD dependent, providing a potential target for breeders.
Abstract
The orientation of plant roots responds to gravity and influences nutrient acquisition. Here the authors show that the formin RMD buffers movement of specialized gravity-sensing organelles and report enhanced RMD expression during phosphate deficiency that could alter root angle to improve phosphate uptake.
Related collections
Most cited references36
- Record: found
- Abstract: found
- Article: not found
Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions.
- Record: found
- Abstract: found
- Article: not found
Roots of the Second Green Revolution
- Record: found
- Abstract: found
- Article: not found