- Record: found
- Abstract: found
- Article: found
The Role of Ceramides in Insulin Resistance
Read this article at
Abstract
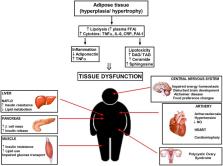
Resistance to insulin is a pathophysiological state related to the decreased response of peripheral tissues to the insulin action, hyperinsulinemia and raised blood glucose levels caused by increased hepatic glucose outflow. All the above precede the onset of full-blown type 2 diabetes. According to the World Health Organization (WHO), in 2016 more than 1.9 billion people over 18 years of age were overweight and about 600 million were obese. Currently, the primary hypothesis explaining the probability of occurrence of insulin resistance assigns a fundamental role of lipids accumulation in adipocytes or nonadipose tissue (muscle, liver) and the locally developing chronic inflammation caused by adipocytes hypertrophy. However, the major molecular pathways are unknown. The sphingolipid ceramide is the main culprit that combines a plethora of nutrients (e.g., saturated fatty acids) and inflammatory cytokines (e.g., TNFα) to the progression of insulin resistance. The accumulation of sphingolipid ceramide in tissues of obese humans, rodents and Western-diet non-human primates is in line with diabetes, hypertension, cardiac failure or atherosclerosis. In hypertrophied adipose tissue, after adipocytes excel their storage capacity, neutral lipids begin to accumulate in nonadipose tissues, inducing organ dysfunction. Furthermore, obesity is closely related to the development of chronic inflammation and the release of cytokines directly from adipocytes or from macrophages that infiltrate adipose tissue. Enzymes taking part in ceramide metabolism are potential therapeutic targets to manipulate sphingolipids content in tissues, either by inhibition of their synthesis or through stimulation of ceramides degradation. In this review, we will evaluate the mechanisms responsible for the development of insulin resistance and possible therapeutic perspectives.
Related collections
Most cited references82
- Record: found
- Abstract: found
- Article: not found
Monocyte chemoattractant protein 1 in obesity and insulin resistance.
- Record: found
- Abstract: found
- Article: not found
An overview of sphingolipid metabolism: from synthesis to breakdown.
- Record: found
- Abstract: found
- Article: not found