- Record: found
- Abstract: found
- Article: found
Interaction of HP1 and Brg1/Brm with the Globular Domain of Histone H3 Is Required for HP1-Mediated Repression
Read this article at
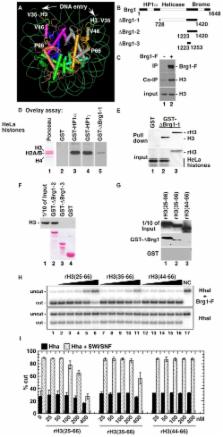
Abstract
The heterochromatin-enriched HP1 proteins play a critical role in regulation of transcription. These proteins contain two related domains known as the chromo- and the chromoshadow-domain. The chromo-domain binds histone H3 tails methylated on lysine 9. However, in vivo and in vitro experiments have shown that the affinity of HP1 proteins to native methylated chromatin is relatively poor and that the opening of chromatin occurring during DNA replication facilitates their binding to nucleosomes. These observations prompted us to investigate whether HP1 proteins have additional histone binding activities, envisioning also affinity for regions potentially occluded by the nucleosome structure. We find that the chromoshadow-domain interacts with histone H3 in a region located partially inside the nucleosomal barrel at the entry/exit point of the nucleosome. Interestingly, this region is also contacted by the catalytic subunits of the human SWI/SNF complex. In vitro, efficient SWI/SNF remodeling requires this contact and is inhibited in the presence of HP1 proteins. The antagonism between SWI/SNF and HP1 proteins is also observed in vivo on a series of interferon-regulated genes. Finally, we show that SWI/SNF activity favors loading of HP1 proteins to chromatin both in vivo and in vitro. Altogether, our data suggest that HP1 chromoshadow-domains can benefit from the opening of nucleosomal structures to bind chromatin and that HP1 proteins use this property to detect and arrest unwanted chromatin remodeling.
Author Summary
HP1 proteins are transcriptional regulators frequently associated with gene silencing, a phenomenon involving masking of promoter DNA by dense chromatin. Owing to their chromo-domain, these proteins can read and bind an epigenetic mark that on many non-expressed genes is present on histone H3 at the surface of the nucleosome (the fundamental packing unit of chromatin). However, the binding to this mark does not explain the repressing activity of HP1 proteins. Here, we show that these proteins can establish a second contact with histone H3, independently of the epigenetic mark. This second contact site is located inside the nucleosome, in a position likely to be inaccessible. Interestingly, this site is also contacted by a subunit of the SWI/SNF complex and this contact is required for the ATP-dependent chromatin remodeling catalyzed by SWI/SNF. We provide evidence suggesting that HP1 proteins use the SWI/SNF chromatin remodeling to gain access to the contact site inside the nucleosome and to prevent further remodeling by competing with SWI/SNF for binding at this position. These observations lead us to suggest that HP1 proteins function as gatekeepers on promoters, detecting and stopping unwanted exposure of internal nucleosomal sites.
Related collections
Most cited references48
- Record: found
- Abstract: found
- Article: not found
Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain.
- Record: found
- Abstract: found
- Article: not found
Chromatin compaction by a polycomb group protein complex.
- Record: found
- Abstract: found
- Article: not found